The source-page: http://carambola.usc.edu/research/biophysics/BacterialFronts.html
Alison Kraigsley ve Paul D. Ronney
Hava-Uzay ve Makine Mühendisliği Bölümü
Güney Kaliforniya Üniversitesi, Los Angeles, Kaliforniya 90089-1453
Steven E. Finkel
Biyolojik Bilimler Bölümü
Güney Kaliforniya Üniversitesi, Los Angeles, Kaliforniya 90089-1340
Soyut
Alevler, polimerizasyon işlemleri ve bazı sulu reaksiyonlar gibi birçok kimyasal reaksiyona giren sistemde meydana gelen kendiliğinden yayılan reaksiyon difüzyon cepheleri hakkında çok şey bilinmekle birlikte, bu engin bilgi tabanı daha önce yayılan hareketli bakteriler gibi biyolojik sistemlere sistematik olarak uygulanmamıştır. Bu çalışmanın amacı (1) bu bilginin biyolojik sistemlere uygulanıp uygulanamayacağını belirlemek ve (2) eğer öyleyse, kendi kendine yayılan cepheler sergileyen biyolojik sistemlerin daha kantitatif ve tahmin edici bir şekilde anlaşılmasını sağlamaktır. Yayılan hareketli bakteriler ve diğer kendiliğinden yayılan cepheler arasındaki analoji, bu cephelerin dinamik özelliklerine göre takip edilecektir. E. coli bakterisini kullanan ilk deneyler gerçekte reaksiyon difüzyon sistemlerine benzer davranışlar göstermektedir. Yayılma oranları, ön eğrilik etkileri, su verme limitleri, stabilite limitleri, yüzdürme etkileri, etkili difüzyon katsayıları ve üreme zaman ölçeği dahil olmak üzere dinamik özellikler üzerine kapsamlı bir çalışma yapmaya çalışıyoruz. Bu bilgi mevcut reaksiyon/difüzyon cephesi analitik/sayısal modellerine girdiler ve belirlenen analojilerin geçerliliği olarak kullanılacaktır. Geleneksel reaksiyon difüzyon sistemleri ve mikrobiyolojik sistemler arasındaki farklar da değerlendirilecektir. Önemli bir fark, bakterilerin evrilmesi ve strese cevap vermesidir; bu, yukarıdaki testi, yaklaşık su verme koşullarında hayatta kalan bakterilerle tekrarlayarak incelenecek bir faktördür. Bu veriler daha sonra, olumsuz koşullar altında bakterilerin hayatta kalma ve adaptasyonunu daha iyi anlamak için kullanılacaktır.
Giriş
Kendi kendine ilerleyen Reaksiyon cepheleri alev, serbest radikal başlatılan polimerizasyon işlemleri ve bazı sulu reaksiyonlar dahil olmak üzere birçok kimyasal ve fiziksel sistemler meydana gelmektedir. Bütün bu sistemler, iki temel özellikleri ile karakterize edilir: aynı zamanda, örneğin, termal enerji için reaksiyonu (hızlandırır reaksiyonun bir ürünüdür (örneğin, alev durumunda bir yakıt-hava karışımı) reaktif bir orta ve bir otokatalist alevlerin vaka). Otokatalist reaksiyonunu başlatmak ve daha otokatalist oluşturma, reaktif ortamına nüfuz zaman Kendi kendine çoğalma meydana gelir. Bu sağlayan reaksiyon-difüzyon herhangi başlatma sitesinden uzakta sabit fiyatla yaymak için cephelerini.
Kendi kendine yayılan bir tepkime-yayılma cephede önemli özelliklerinden ikisi, çoğalma oranı (ler) ve su verme sınırı. Yayılma oranı genellikle toplam reaksiyon zaman ölçeği (τ), otokatalist difüzyon katsayısı (D) bağlı reaktif ortama ön gelişmeler biçimi ilişkisini s ≈ (D/τ)1/2 ‘e göre olan hızıdır. bu da t reaktanların konsantrasyonuna ve etkinliğine bağlıdır. Söndürme sınırı bir parametrenin minimum veya maksimum değer (örneğin, en az reaktif konsantrasyonu veya kanal genişliği) olan bir sürekli yayılan ön mevcut olabilir. Alevler kimyasal reaksiyonlar ve difüzif ve konvektif taşıma arasında meydana gelen kavrama [[1]] Polimerizasyon cepheleri [[2]] ve sulu reaksiyonları [[3]] de nicel olarak anlaşılmaktadır.
Şaşırtıcı bir şekilde, sorgulama analog hattı ilişkisi s ≈ (D/τ)1/2 giden, daha reaktif yayan bir cephede ilk modelleme da, mikrobiyolojik sistemlere uygulanmamıştır bağlamında 1937 yürütülmüştür, bulaşıcı hastalıkların yayılması (bakınız [[4]]). Araştırmamızın amacı bunu yaparken fizibilitesini değerlendirmek ve bu tür mikrobiyolojik sistemlerin daha nicel bir anlayış geliştirmektir. Çoğu mikrobiyolojik çalışmalar bireysel bakteri ve bireylerin davranışlarını sayma odaklanır. Bu çalışmada termodinamik yasaları uygulamak ve bireylerin çok sayıda toplu davranışlarını incelemek. Bu çalışma için odaklanmayı tercih test durumu çok yaygın ve çok çalışılan bir Escherichia coli bakteri, (diğerleri gibi) kendi dokunaçlarını kullanılarak veya yüzer bir hareketli bakteri kamçı sıklıkla daha yüksek besin konsantrasyonu bölgeleri aramaya yön değiştirirken onun besleyici ortam içinde. Özellikle bakteri davranışı iki modu vardır: kendi kamçı bunun nedeni, flagella küçük bir net hareket yönü değiştirmek burada moduna daha çok ya da daha az düz çizgi halinde itmek ve “kurutma” için döndüğü “çalıştırma” modunda [[5]]. (Bakteri hareketi bakteri arasındaki elastik çarpışmalar ilişkili değildir molekülünden farklı olarak, mekanizmanın tamamen farklı olmakla birlikte, çünkü) elde edilen hareket klasik Fickian difüzyon neden moleküllerin rastgele yürüyüş biraz benzer. Daha yüksek D ile sonuçlanır çakılır daha elverişsiz gradyanlannda, bu yeni yol bulmak için izin alt D [sonuçlanan, daha çakılır ise aynı zamanda, uygun bir besin gradyanı içinde, bakteri, daha çalışır Hata! Bilinmeyen anahtarı argüman.]; bu da artmış besin seviyelerinin bölgelere göçün lehinedir yanlı rasgele yürüyüş yol açar.
Hareketli bakterilerin bu özellikleri, reaksiyon difüzyon cephede olarak tarif etmede kullanılabilir. Yayılma hızı (ler), E. coli, bir% 0.3 agar ortamında, ölçülen (Ön Sonuçlar bölümüne bakınız) yaklaşık 4.5 mm/saattir. Üreme zaman ölçeği (τ), E. coli 20 dk Hata! Bilinmeyen anahtarı argüman. Yana yayılma hızı, s ≈ (D/τ)1/2 , D ≈ s2τ, böylece D ≈ 1.5 x 10-5 cm2/sn. Bu değer, D parçacık tarafından çarpılan bir parçacık hızı (c) serbest bir yol (λ), ortalama orantılı olduğunu göstermektedir moleküllerin kinetik teorisine göre beklenen değer ile tutarlıdır yani. Molekül parçacıkları birbirleriyle çarpışmadan veya hareketli bakteriler durumunda, yön değiştirme öncesinde hareket mesafesi durumunda. l bakteri miştir yüzmek zaman (τ) ile çarpılır C olarak tahmin edilebilir. İçin, E. coli, 30 saniye için bir yönde 21 değişikliklerle 21 µm/s arasında bir ortalama yüzme hızı, ölçülmüştür [Hata! Bilinmeyen anahtarı argüman.]. Bu ≈ 1.4 s ve λ ≈ 3.0 x 10-3 cm ve böylece etkin difüzyon D ≈ 6.3 x 10-6 cm2 D değeri aynı büyüklük düzeyindedir D ≈ s2τ.
Biz reaksiyon-difüzyon cephede olarak hareketli bakterilerin yayılmasını karakterize etmek makul olduğu sonucuna vardık. Birçok mikrobiyolojik çalışmalar sıcaklığında ya da sistem üzerinde uygulanır besin gradyanı sistemin tepkisini ölçmek; Biz bakteriler kendi geçişlerini üretebilir ve böylece kendiliğinden yayılır olabileceğini göstermek istiyoruz. Tablo 1 alevler arasındaki önerilen analoji (tipik reaksiyon difüzyon ön) ve mikrobiyolojik sistemleri göstermektedir.
| Alev veya moleküler özellik | Mikrobiyolojik eşdeğer |
| Sıcaklık | Bakteri konsantrasyonu |
| Yakıt | Besinler |
| Isı yayınım ≈ cλ | Bakterilerin yayılımı |
| Yakıt yayınım | besin Difüzivite |
| Ses hızı (c) | “Koşmak” modunda bakteri hızını Yüzme |
| Serbest yolu (λ) ‘ne | Ortalama zaman çarpılacak c arka modunu takla çalışma moduna geçmek ve |
| Reaksiyon zaman ölçeği (τ) | Üreme zamanı |
| Isı kaybı | (Bireysel bakterinin) Ölüm |
| su verme | Ölüm (bütün bakteriler) |
| Tablo 1. Alevler ve mikrobiyolojik cepheler arasında önerilen analoji | |
Alevler ve hareketli bakterilerdeki karmaşık bir faktör, etkili difüzyon katsayısının ürün konsantrasyonuyla birlikte değişmesidir. Alev durumunda, ürünlerin üretimi, neredeyse tüm durumlarda D’yi artıran daha yüksek sıcaklığa yol açar. Hareketli bakteri durumunda, besin konsantrasyonunun yüksek olduğu durumlarda bakterilerin “çalışma” modunda daha az zaman harcadıkları ve “tambur” modunda daha fazla zaman harcadıkları iyi bilinmektedir (neden çimlerin yeşil olduğu zaman etrafa bakmalısınız?) Bu ortalama serbest yolun l daha düşük olacağı ve dolayısıyla besin konsantrasyonu yüksek olduğunda etkili difüzyonun daha düşük olacağı anlamına gelir. Yayılma ön cephesinde ön cephenin ön kenarındaki az sayıda bakteri yüksek besin konsantrasyonları görür ve bu nedenle düşük D değerine sahiptir. Ön cephenin arka kenarındaki yüksek bakteri konsantrasyonları (reaksiyonun “ürünü”) daha düşük besin konsantrasyonuna sahip bir ortam ve bu nedenle “daha fazla” çalışacak ve “daha az” takla “kalacaktır, l artacak ve böylece D artacaktır. Dolayısıyla, alevlerle yapılan benzetme oldukça geniştir.
Ön sonuçlar
Yukarıdaki tartışma bizim önermeler uygulanabilirliğini test etmek ön deneyler için teşvik etti. Bu deneyler, (aksi belirtilmediği sürece)% 1 NaCl,% 1 tripton,% 0.5 maya ve% 0.1 agar içeren bir su bazlı ortam maddesinin kullanımı ile standart bir petri levhalar içinde 37 ° C’da gerçekleştirilmiştir. Agar ortamı viskozitesini arttırır ve esas olarak herhangi bir yığın hareketi engeller. Ortam (genellikle çanak merkezinde) bir noktada bakteri ve bakteri ön görsel olarak gözlemlenmiştir sonuçtaki önceden (varsa) ile aşılandı. Birçok suşu E. coli test edilmiştir. Bir ilk geçici sonra, her zaman ön yan çapı olan bir lineer bir artış sergilemiştir. Bir örnek Şekil l’de gösterilmiştir. 1.
Şekil 1, ön yarıçapı küçüktür ve ön kavis yüksek, ön başlangıçta daha yavaş ilerler göstermektedir. Diğer tüm testler aynı eğilimi gösterdi. Bu davranış, (otokatalitik ürün tepkime maddesi daha düşük bir difüzyon sahip olduğu kimyasal cephede görülmektedir , yani alev, yüksek termal yayılma ve düşük yakıt yayılma durumunda.) Ve nedenleri de [anlaşılmaktadır [6]]. Tripton besin yayılma benzer moleküllerle benzer şekilde, bilinmemekle birlikte 10-7 cm2/s değeri tahmin edilebilir, görünen yayılma daha düşük olan, E. coli Yukarıda tahmin olarak. Sonuç olarak, Şekil. 1 ‘de gösterilen davranış ön çoğalma reaksiyonu difüzyon modeli ile tutarlıdır. Aynca, alev teori [Hata! Bilinmeyen anahtarı argüman.] Difüzyon reaktifinin ürünün oranı çok değeri 1 farklı olarak, ya daha yüksek ya da daha düşük olması durumunda, ön kararsızlıklar çeşitli neden olabileceğini göstermektedir. Mikrobiyolojik sistemlerde desen oluşumu iyi bilinmektedir [[7]], ancak reaksiyon difüzyon teorisi ölçmek ve bu gibi desenler tahmin etmek için kullanılır edilmemiştir. Biz Agar konsantrasyon yoluyla orta yayılma gücünü değiştirerek bu tür kararsızlıkları araştıracaktır. Ayrıca, şüpheli çalıştırma çamaşır zamanlama bu seçim E. coli Bu D etkilediği için tesadüfi değildir; D çok düşük veya çok yüksek ise, istikrarsızlıklar bu hayatta kalmak için zararlı olabilir sonuçlanır. Biz hızına Agar konsantrasyonunun etkisini inceleyerek bu kavramı takip ve mikroskop altında bakteri zamanlamasını kırana çalıştırın ve (ya anında ya da evrimsel süreçler tarafından) bakteri uyarlar olmadığını belirlemek hızı ve zamanlama kırana çalışacaktır kararlı cephelerini muhafaza edilmesi için etkili bir D değiştirmek için kullanılır.
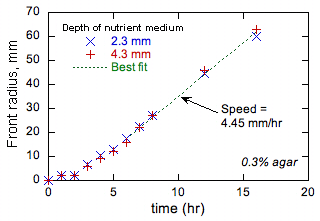
Şekil 1. Zamanın bir fonksiyonu olarak bakteri ön yarıçapı üzerindeki deneysel veriler.
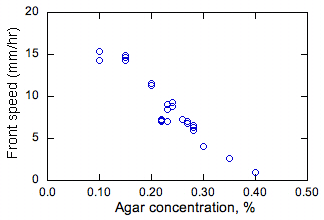
Şekil 2. Agar ve çözelti derinliğinin bakteri cephelerinin sabit yayılma hızları üzerindeki ölçülen etkisi.
Agar konsantrasyonunun etkisi ve yayılma hızının s sabit değerleri besleyici ortam derinliği Şekil l’de gösterilmiştir. 2.% 0.5 veya daha büyük konsantrasyonda agar (gösterilmemiştir), ortam esas itibarıyla sağlam ve bakteriler sadece yayılan olup toplu olarak orta yüzeyi üzerinde. Bakterilerin Difüzyon benzeri davranış onlar yüzemem çünkü bu durumda olası değildir. % 0.4 ve daha düşük konsantrasyonda agar, bakteri vasıtasıyla yayılır. En yüksek yayılma hızları (ler) olan en düşük agar konsantrasyonlarının varlığında (ve ortamın bu şekilde düşük viskozite) gözlenmiştir. Düşük viskozitede, bakteri hızlı yüzebilir ve böylece daha yüksek etkili D olması gerekir, (ler ile orantılı olduğu için daha yüksek s giden bu durum beklenen D1/2 yukarıda tartışıldığı gibi).
Çözeltinin derinliği s üzerinde neredeyse hiçbir etkisi olduğu bulunmuştur. ön kısmi yayılım sonrasında orta enine kesitleri alarak, bu kapaklar genelde dikey boyutta tek biçimli olduğu bulunmuştur. Böylece yüzdürme akımı dikey yönde çözelti s derinlemesine veya düzgün olmayan, aksi takdirde bir etki, bu kapaklar etki değildir, beklenir. Testler havada gerçekleştirildi olduğundan, bu gözlemler de yüzeyin altında farklı olurdu, ortama oksijen difüzyon önemli olmadığını besin solüsyonunun yüzeye yakın aksi büyümesini gösteriyor. Bu son sonuç özelliği bakterilerin büyüyen olduğunu göstermektedir.
“Söndürme” sınırları bakteriyel cephelerde mevcut olup olmadığını belirlemek için, bir deney yapılmıştır, ki burada bakteri cephede antibiyotik duvarlı, geniş ve dar kanallar yayılması çalıştı. Antibiyotikler otokatalitik ürün (bakteriler) çıkarmak için, duvar etkisi alevler soğuk duvarlara ısı kaybıyla benzer. Dar kanallarda ön yayılımı için, iyi bilinen [olduğunu Hata! Bilinmeyen anahtarı argüman.] W kanal genişliği ve D, yayıcılığıdır Peclet sayısı Pe = sw/D, kritik değerden daha düşük olduğunda, bir söndürme sınır kanal duvarlarına kayıp ısı nedeniyle oluşur. Bu nedenle, ısı kayıplarının varlığında, cephe geniş kanallar (yüksek Pe), ancak dar kanallara (düşük Pe) ile ilerleyebilir. Şekil 3, cephede bir söndürme sınırı gösteren, geniş kanal değil, dar kanaldan yayıldığını göstermektedir. söndürme ilişki uygulanabilirliği Pe = Sw/limit sabit bu çalışmada test edilecektir = d.
Tripton besin konsantrasyonunun etkisi de test edilmiştir; s nominal değerinden tripton konsantrasyonunun iki katına neredeyse değişmeden oysa, yarı nominal değerden bir yayılma meydana getirilebileceği bulunmuştur. Bu da, antibiyotik duvarların yokluğunda bir söndürme sınırı gösterebilir. Yakıt konsantrasyonu çok düşük olduğunda benzer davranışlar nedeniyle radyasyonlu ısı kayıpları alevler içinde gerçekleşir. Bu duvar içermeyen sınırı araştırmak ve düşük besleyici seviyelerinin ön yayılmasını bastırmak için sorumlu kayıp mekanizmasını belirlemek için çalışacaktır.
Elde etmek için bakteri ortamının Titering sürekli, birincisi arkasında, daha sonraki zamanlarda, ikinci bir tepe ön arkasına önemli bir düşüşü ile takip gelen ön bakteri konsantrasyonu önemli bir tepe gösterdi ve sayımları. Bu belirlemek için biz analizini titering kullanacağız içinde Şek. 3. Gelecekteki çalışmada gösterilen görüntülerle tutarlı ön kalınlığı ≈ D/s teorik tahmin edildiği gibi [eğer Hata! Bilinmeyen anahtarı argüman.]. Şekil görsel inceleme üzerine dayanmaktadır. 3 ön kalınlığı tahmini D/s = (1.5 x 10 civarındadır, yaklaşık 1-3 mm, mm2/sec)/(4.5 mm/h) = 1,2 mm. bakteri vahşi suş ile elde edilen sonuçlar bir miktar daha hızlı ilk geçici ve genetik olarak işaretlenmiş türler ile karşılaştırıldığında sürekli sabit bir yayılma hızına ulaşmak için daha kısa bir zaman ve mesafe gösterdi.
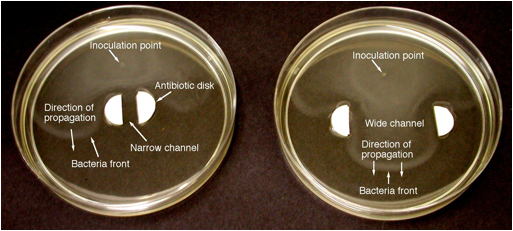
Şekil 3. Dar (6 mm, sol) ve geniş (35 mm, sağ) kanallardan Kanamisin antibiyotik yan duvarları (yan başına 100 mikrolitre Kanamisin) ile yayılan hareketli E. coli bakteri önlerinin fotoğrafları, aşılamadan 6.5 saat sonra alındı. Her iki durumda: 2086 E. coli’nin yabani türü,% 0.1 ağar, standart besin maddesi.
Güncel araştırma programı
Bu ön deneysel sonuçlarda tarif edilen reaksiyon difüzyon modelinin görünür başarısına dayanarak, aşağıdaki araştırma programını takip ediyoruz:
- Çoğalma oranları belirlemek E.coli besin konsantrasyonu, (ağar konsantrasyonunun değiştirilmesi yoluyla) orta motilite, ortam sıcaklığında, ortam oksijen konsantrasyonu (aerobik genel anaerobik ortam) bir fonksiyonu ve besleyici ortam tabakasının kalınlığı cephede. Bu testler, yukarıda tarif edilen ön-deneylerde ile aynı şekilde gerçekleştirilir. Buna ek olarak, cephede iyi görülmesi, kullanacağımız E. coli Molecular Probes, Inc., (Eugene, OR) ile ilgili belirli bir floresan boyalar. Cephede aralığının daha iyi görülmesi. Bir ultraviyole ışık kaynağı (satın alınmış) ya da Ar-iyon lazeri floresan boyalar uyarmak için kullanılır.
- İlişkisi D ≈ s2τ göre D belirlemek ton ile karşılaştırmak ilişkiye dayanarak beklenen D = c2τ yönünü değiştirmek için ortalama zaman t hızı (c) ve (Video mikroskopisi kullanılmıştır) ölçülür t. Çalışma çamaşır zaman ölçeği tipik olarak 1 sn ve video çerçeve hızı saniyede 30 olduğu Standart video yeterlidir.
- Söndürme sınırlarını belirlemek ve genişliğini (w) sahip olan değişken kanallar yayılması cepheleri izin vererek Peclet sayısının sınırlanması antibiyotik duvarlar (Kanamisin veya benzeri) ve Pe sınırı, sabit olup olmadığını belirlemek, yani, yayılma hızı (ler) de olduğu 1/w ile orantılı sınır?
- (Konsantrasyon agar değiştirilmesi için, besin konsantrasyonunun azaltılmasıyla) duvar yokluğunda söndürme sınırını belirler ve zarar mekanizması (tespit yani. Hücre ölümü mekanizması) bu sınırın sorumlu değildir.
- Besleyici ortam ve besin konsantrasyonu konsantrasyonda agar değiştirilmesiyle (besin) difüzyon reaktifinin ürün (bakteri) oranının bir fonksiyonu olarak denge sınırlarını belirler. Floresan boyalar düzgün olmayan cephede ve desen oluşumu görüntüleme kolaylaştıracaktır.
- Ön çoğalması üzerinde yüzdürme etkisi olup olmadığını belirlemek. Bu yana, E. coli bakteri düşük viskoziteli suda biraz farklı bir yoğunluğa sahip bir yüzdürme etkisi ortaya çıkar. Bir çok araştırmacı her türlü sıvı akışını önlemek için bir yüksek viskoziteli agar ortamını kullanmak, ama birçok gerçek E.coli ortamları su benzeri viskoziteye sahiptir. Yüzdürme etkileri dikey paralel plaka (akışkanlar mekaniği literatüründe “Bütün-Shaw” hücre olarak da adlandırılır) aparatı ve ön deseni ve yayılma hızının gözlemlenecektir agar-içermeyen ortam içinde ön yayılma test edilmesi ile incelenecektir.
- Reaksiyon-difüzyon cephede oluşan mevcut analitik/sayısal modeller kullanılarak bu sonuçları Model.
Tabii ki, geleneksel reaksiyon difüzyon sistemleri ve mikrobiyolojik sistemler arasında da bazı farklılıklar vardır. Önemli bir fark, bakterilerin evrilmesi ve strese cevap vermesidir; bu, yukarıdaki testi, söndürme koşullarında hayatta kalan “deneyimli” bakterilerle tekrarlayarak çalışacak bir etkendir. Yukarıda tartışıldığı gibi başka bir fark, yanlı difüzyon özellikleridir. Bu önyargı, Fick Yasası’nın yerine doğrusal olmayan bir yayılma ilişkisi kullanılarak modellenebilir.
Pratik önemi
Bu çalışma, hareketli bakterileri içeren yeni bir sınıf cephelerin dinamiklerini inceleyen bir çalışmadır. Ön dinamiklerin incelenmesi yukarıda tartışıldığı gibi birçok uygulamaya sahiptir. Bununla birlikte, bu çalışmalar mikrobiyologlar için özellikle ilgi çekicidir, çünkü uzun süreli hayatta kalma ve bakterilerin kullandığı evrim mekanizmaları ile ilgilidir. Bu bakteriyel sistemlerdeki evrimsel değişim oranlarını etkileyen ana faktörlerden biri çevresel stresin yanı sıra bir ortamdan diğerine geçiş seviyeleridir. Geliştirmekte olduğumuz kültür sistemleri, bu gelişim ve hayatta kalma sorularının ele alınmadığı yeni ortamları temsil ediyor. Bu çalışma, bildiğimiz kadarıyla bu etkileri biyofiziksel olarak karakterize etmek için ilk girişimi sağlıyor.
Anaerobik koşullar altında büyüyen E. coli‘nin çalışması özellikle ilgi çekicidir. E. coli (ve tüm diğer enterik bakteriler) insan sindirim sistemi ile birlikte büyüyünce, anaerobik koşullar altında büyürler. Bu bakterinin çok çeşitli çevresel nişlerde hayata nasıl adapte olduğu konusundaki genel anlayışımızı daha da genişletmek için oksijensiz ortamda bakteriyel stres yanıtlarını incelemeye başlamak istiyoruz. Örneğin, şu anda, DNA’nın mekanizmalarının Prof. Finkel’in araştırdığı (ve yeni ortamlara adaptasyon için önemli olduğunu gösterdiği) anaerobik koşullar altında çalıştığı onarım ve mutasyona zarar verip vermediği henüz bilinmiyor. Bu çalışmalar, temel araştırmalardan farkı daha tıbbi olarak ilgili alanlara köprü kılacaktır.
Referanslar
[1]. Williams, F. A., Combustion Theory, 2nd Ed., Benjamin-Cummins, 1985.
[2]. Pojman, J. A., Hyashenko, V. M., Khan, A. M., “Free-radical frontal polymerization: self-propagating reaction waves.” J. Chem. Soc., Faraday Trans. 92, 2825 (1996).
[3]. Epstein, I. R. Pojman, J. A. An introduction to nonlinear chemical dynamics, Oxford, 1998.
[4]. Winfree, A.T., The Geometry of Biological Time, Springer-Verlag, 1990; Murray, J.D., Mathematical Biology, Springer-Verlag, 1993.
[5]. Berg, H. C., “Motile Behavior of Bacteria” Phys. Today 53, 24 (2000).
[6]. Lewis, B., von Elbe, G., Combustion, Flames, and Explosions of Gases, 3rd ed., Academic Press, 1987.
[7]. Budrene E.O., Berg H. C., “Complex patterns formed by motile cells of E. coli,” Nature 349, 630 (1991).